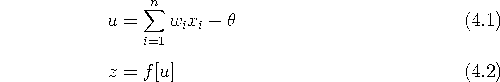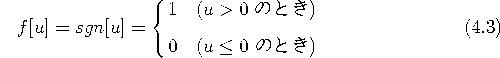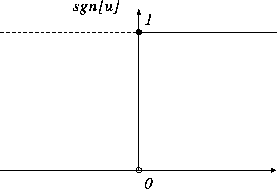神経細胞のモデル化の研究には、大きく分けて、神経細胞自体に注目し、神経細胞内におけるインパルスの発生や伝搬の機構をモデル化する立場と、神経細胞の微細な構造にはとらわれずに神経回路網を構成している要素としてモデル化する立場とがある。
前者の研究として代表的なものに、神経線維の膜電位の変化を記述するHodgkin-Huxleyの方程式(Hodgkin-Huxley,1952)がある。
この方程式は、「細胞膜上の Na ![]() チャネル、K
チャネル、K ![]() チャネルの透過性は膜電位によって変化する」というモデルに基づいて作られた方程式で、膜を横切って流れる電流を、膜電位およびその時間微分の関数として表したものである。
この方程式を、神経線維の伝送路としての特性を表す方程式と組み合わせることによって、神経線維における電気的現象を記述することができる。
神経線維において観察される現象は、Hodgkin-Huxley 方程式によってほぼ完全に説明することができるが、この方程式は非線形微分方程式であるため取り扱いが難しく、その解の振舞いについては完全には分かっていない。
なお、これを単純化したものに Fitzhugh-Nagumo の方程式がある。
チャネルの透過性は膜電位によって変化する」というモデルに基づいて作られた方程式で、膜を横切って流れる電流を、膜電位およびその時間微分の関数として表したものである。
この方程式を、神経線維の伝送路としての特性を表す方程式と組み合わせることによって、神経線維における電気的現象を記述することができる。
神経線維において観察される現象は、Hodgkin-Huxley 方程式によってほぼ完全に説明することができるが、この方程式は非線形微分方程式であるため取り扱いが難しく、その解の振舞いについては完全には分かっていない。
なお、これを単純化したものに Fitzhugh-Nagumo の方程式がある。
後者の立場では、1つ1つの神経細胞の機能の記述は単純にして、細胞をつなぎ合わせたときに全体で発揮される機能に着目する。 以下は、この立場から神経細胞のモデル化について詳しく説明する。
神経細胞の機能をごく単純化してとらえると、つぎのようにまとめることができる。
まず、神経細胞は多くの細胞から入力を受けて出力を出すことから、多入力1出力の素子であると考えてよい。 他の細胞の出力はシナプス結合を介して入力として与えられるが、このとき、シナプス前細胞がシナプス後細胞へ及ぼす影響の強さは各細胞によって異なると考えられる。
いま、1つの神経細胞が n 個の細胞から入力を受けているとする。
シナプス前細胞の出力を ![]() 、それぞれのシナプス結合の強さを
、それぞれのシナプス結合の強さを ![]() と表すと、最も単純には、 i 番目の細胞からの影響は
と表すと、最も単純には、 i 番目の細胞からの影響は ![]() と表される。
この
と表される。
この ![]() をシナプス荷重、またはシナプス伝達効率とよぶ。
各細胞からの影響
をシナプス荷重、またはシナプス伝達効率とよぶ。
各細胞からの影響 ![]() が単純に加算されたものが細胞の入力になるとすると、この和が閾値
が単純に加算されたものが細胞の入力になるとすると、この和が閾値 ![]() を越えた時に、細胞が興奮する。
さらに、軸索は減衰のない伝送路であると仮定すれば、このようにして生じた興奮がその細胞の出力になると考えられる。
を越えた時に、細胞が興奮する。
さらに、軸索は減衰のない伝送路であると仮定すれば、このようにして生じた興奮がその細胞の出力になると考えられる。
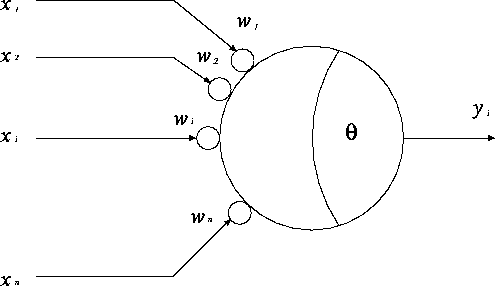
以上の説明から図4.1のようなモデルとなり、また、このモデルを式で表すと次のようにまとめられる。
式(4.1)は、ほかの細胞からの荷重和が細胞に与えられ、それから閾値を引いたものが u であることを意味している。 この u を膜電位、もしくは、内部ポテンシャルとよぶ。 式(4.2)は、細胞の出力は膜電位 u の関数として決まることを表している。 f[u] は出力関数とよばれる関数で、一般に単調非減少関数である。
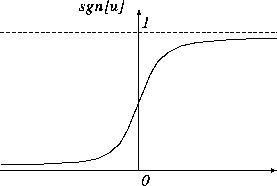
入力 ![]() 、出力 z のとりうる値としては、0と1や -1 と1といった2値のみをとる2値モデルと、図4.2のようなシグモイド(S字形)関数、線形関数などをとる連続値モデルがあるが、ここでは、取扱いが簡単なことから2値モデルについて考える。
2値モデルの場合、 f[u] は式(4.3)で定義され、図で示すと、図4.3のような階段関数となる。
、出力 z のとりうる値としては、0と1や -1 と1といった2値のみをとる2値モデルと、図4.2のようなシグモイド(S字形)関数、線形関数などをとる連続値モデルがあるが、ここでは、取扱いが簡単なことから2値モデルについて考える。
2値モデルの場合、 f[u] は式(4.3)で定義され、図で示すと、図4.3のような階段関数となる。
このように、出力として0、1の2値をとるようなモデルを、入力の線形和( ![]() )が閾値
)が閾値 ![]() を越えたときのみ1を出力することから、特に``線形閾値素子''モデルという。
を越えたときのみ1を出力することから、特に``線形閾値素子''モデルという。